I’m very pleased to present another UALVP-related study today, this time by Caleb Brown (formerly at the University of Calgary and now at the University of Toronto). Caleb recently published a paper in PLoS ONE featuring one of my favourite UALVP specimens, our Stegoceras partial skeleton, UALVP 2.
1. What inspired you to conduct this study?
I was initially interested in pachycephalosaur postcranial anatomy for the purpose of differentiating between isolated pachycephalosaur postcranial material and those of basal ornithopods (like Thescelosaurus and Parksosaurus), on which I was doing my Masters research at the University of Calgary with Anthony Russell. In order to get a better understanding of the postcranial anatomy of these animals I went to the source, Stegoceras – UALVP 002, one of the best (if not the best) pachycephalosaur skeletons known, and the first postcranial skeleton discovered. In addition to other things, I was initially struck by the presence of large numbers of bony elements that I could not identify and that did not match the morphology of other ornithischians.


These elements looked superficially like gastralia, and indeed that is what they were identified in Gilmore’s 1924 description. But ornithischians were not supposed to have gastralia, so my interest was peeked. Investigation into the literature revealed that others had worked on these enigmatic elements; Marya´nska and Osmólska (1974) found similar elements in the tail of Homalocephale in Mongolia, illustrating they were not gastralia, and Sues and Galton (1987) correlated the structures between Homalocephale and Stegoceras. Particularly interesting was the articulated series found in the tail of Homalocephale. These showed a distinctive pattern that matched myomeres and myosepta, the sideways “w” shaped muscles and tendons, seen in fish.
Fortunately, with funding from Lubrizol Corp. and Montessori High School (University Circle, Cleveland, OH) I was able to accompany Michael Ryan (Cleveland Museum of Natural History) and David Evans (Royal Ontario Museum) to Mongolia to do fieldwork in the summer of 2009. I was also fortunate enough to be able to examine the Homalocephale specimen while I was there. This allowed me to test my ideas about the deep homology of these interesting structures.


2. What’s so special about pachycephalosaur tails, anyway?
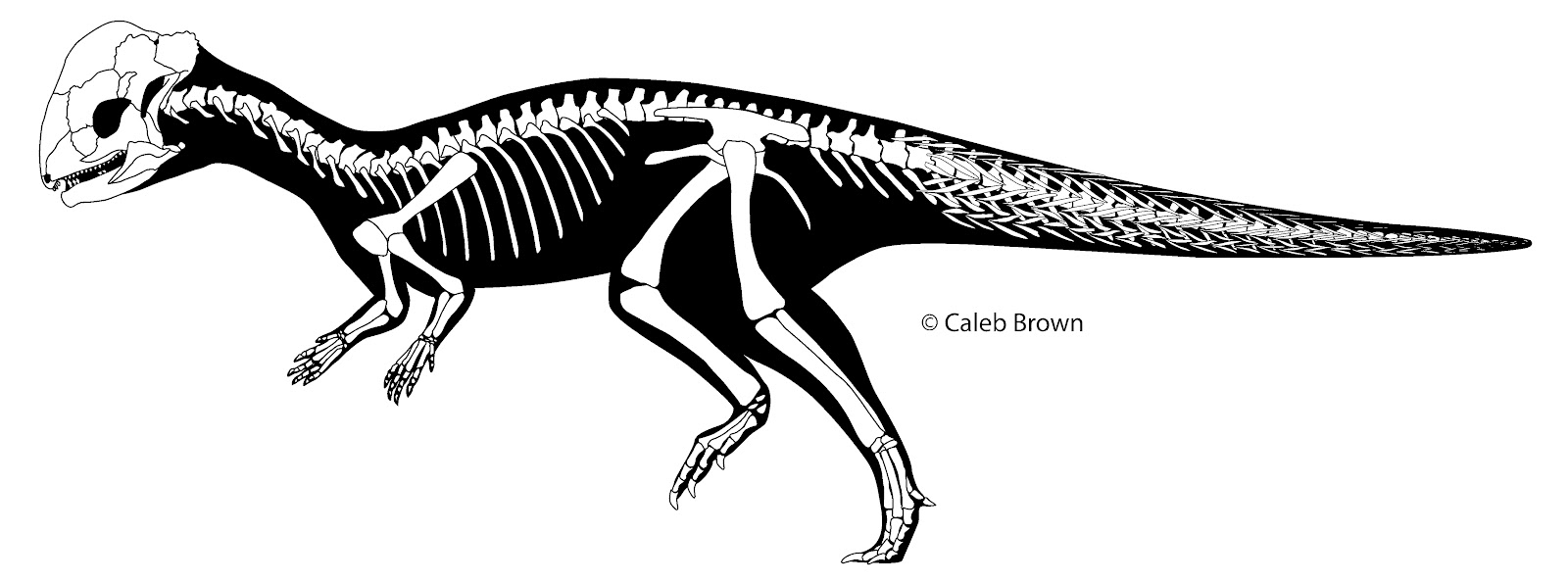
First off, pachycephalosaur tails, like the rest of their postcranial skeletons, are rare. Often with dinosaurs you find the rest of the skeleton but are missing the most important part, the head. This is not true for pachycephalosaurs, which we know almost everything about based on the skull. You can count on one hand the number of partial skeletons known (and these are partial skeletons). What we know about pachycephalosaur skeletons is limited to these few specimens. They are special in that when preserved they show a unique morphology of having a halo of superficial “W” shaped elements forming a cylinder around the entire circumference of the tail. This is not seen in other dinosaur groups, or any other tetrapod. That is not the only odd thing though; they lack the deep longitudinal or paraxial tendons seen in most other ornithischian groups and they have elongated and highly bowed caudal ribs. These three things may be related, but that is not yet clear.
3. What is the difference between gastralia, ossified tendons, and ossified myorhabdoi?
This is an interesting question with a bit of a complex answer. All of these structures are similar in that they are not endochondral bones, that is they do not develop from a cartilaginous precursor, which is the case with the majority of the postcranial bones in most taxa.
Gastralia are dermal or intramembranous bones that are associated with the abdominal musculature, and can be associated with respiration. They were likely the primitive condition for tetrapods but today are restricted to Crocodylia, Sphenodon, and possible the plastron of turtles (Classens, 2004).
The term ‘ossified tendons’ describes a variety of structures including ossified myorhabdoi. Although this term would include any ossification of the connective tissues articulating muscles to bones, its usage in dinosaurs, particularly ornithischians, usually refers to longitudinal paraxial structures along the dorsal or caudal vertebral series. These tendons often have the pattern of either a trellis or longitudinal bundles, can be epaxial or hypaxial, and are usually closely associated with the vertebrae (Organ 2006). Ossified myorhabdoi are restricted to the caudal musculature, and are essentially ossified myosepta. Unlike the majority of the paraxial tendons, these are superficial, forming a halo around the circumference of the tail where the transverse skeletogenous septum intersects with the integument, and preserve a morphology reminiscent of the undifferentiated myoseptal musculature of fish. They are also different in their histological structure (Organ and Adams, 2005). We still know very little about ossified myorhabdoi and hopefully discovery of additional specimens and more research on extant taxa will reveal more regarding their significance.
4. Why don’t other dinosaurs have a caudal basket?
It is often hard to answer why some groups have a structure while other don’t, and this becomes particularly difficult when the function of the structure is not fully understood. If the function of the ‘caudal basket’ is to rigidify the tail of pachycephalosaurs, then the reason that other groups don’t have it is because many have found a different solution to the same problem. Many other ornithischians have longitudinal or paraxial tendons (usually called “ossified tendons”) in the form of a trellis or bundles. Some theropods stiffen their tail by extending the zygapophyses across numerous vertebrae. Until the function of these structures can be better established, we may not know the full significance of their occurrence.

5. Does the presence of a caudal basket tell us anything about head-butting behaviour in pachycephalosaurs?
The caudal basket likely had significant implications for the posture and locomotion of pachycephalosaurs. It has been suggested by previous authors that it helped the tail to act as a tripodal prop, potentially during intraspecific behaviour. It would also have greatly stiffened the tail. Our analysis is consistent with these interpretations, and in that manner is consistent with the idea of head-butting behaviour in pachycephalosaurs.
The presence of the caudal basket has also been used to support the idea of agonistic flank butting behaviour in pachycephalosaurs (Goodwin et al., 1998), with the caudal basket acting as armor. We suggest that the morphology of the myorhabdoi is not consistent with armor seen in other groups, and this function in pachycephalosaurs seems unlikely.
Thanks very much Caleb! You can read more about pachycephalosaur tails in:
Brown CM, Russell AP. 2012. Homology and architecture of the caudal basket of Pachycephalosauria (Dinosauria: Ornithischia): the first occurrence of myorhabdoi in tetrapoda. PLoS ONE 7(1): e30212.
